
Фрисман Е.Я., Жданова О.Л., Павлова E.A.
ИАПУ ДВО РАН, лаборатория математического
моделирования экологических систем
We investigate the fitness differences model with Mendel limited population in cases of linear and exponential rules of selection. It was made the analytical and numerical investigation of the model. The possibility of population size fluctuations with stable genetic structure was showed. Also were found cooperative fluctuations of genetic structure and populations size. This result contradicts to well-known hypothesis of ``genetic control''. We've found stable chaotic attractors with non-integer dimensions.
Динамика популяций и биоценозов может описываться дифференциальными и рекуррентными уравнениями. Существует ряд биологических видов с неперекрывающимися поколениями (многие насекомые, лососевые рыбы и много других), которые естественно описывать, пользуясь дискретными динамическими системами. Дискретные модели простейших биоценозов позволяют описать такие реальные эффекты, как возникновение циклов динамики численности лимитированной популяции, псевдослучайное поведение экосистем и др., наблюдаемые на реальных и лабораторных популяциях [5].
Детерминированные модели - модели, построенные при помощи аппарата дифференциальных или разностных уравнений для частот генотипов или гамет.
Сформулируем предположения, общие для всех детерминированных задач:
Поведение численности популяции во времени - одна из основных характеристик биологического вида. Не существует ни одной популяции, численность которой не испытывала бы изменений. Некоторые из этих изменений носят сезонный характер и связаны с колебаниями климатических условий в течение года, другие затрагивают периоды времени, включающие годы и десятилетия. Колебания численности обладают различной регулярностью и размахом, могут сопровождаться существенными перестройками структуры популяции, приводить к изменениям роли и значимости различных видов в биосистемах. До сих пор нет единой точки зрения на то, какие же факторы являются ведущими, определяющими колебания численности, ее подъемы и спады.
Существует несколько гипотез о конкретных механизмах внутрипопуляционной регуляции уровня численности через изменение плодовитости, смертности, скорости созревания и т.п. Наиболее популярны две из них: гипотеза стресса и гипотеза генетического контроля. Согласно первой, при излишне высоких плотностях населения в пиках численности у животных появляются признаки стресс-синдрома: увеличиваются надпочечники, крайне усиливается агрессивность так, что нормальное размножение почти полностью прекращается и, в связи с этим, падает рождаемость и растет смертность. В рамках второй гипотезы колебания численности популяции связывают с изменением их генетического состава. Заметим, что в рамках первой гипотезы остается непонятен эволюционный механизм, закрепивший резкие закономерные снижения приспособленности в пиках численности. Вторая же гипотеза вступает в противоречие с предположением, что наличие плотносто зависимого отбора является скорее всего следствием колебания численности, а не их причиной, так как такой отбор может появляться только в флюктуирующих популяциях [5].
Для принципиального понимания основных результатов, полученных при
исследовании действия отбора в рамках детерминистических динамических
моделей, достаточно рассмотреть весьма простую ситуацию, когда все
адаптивное разнообразие в популяции определяется одним диаллельным
локусом ![]() с аллеломорфами
с аллеломорфами ![]() и
и ![]() , причем фенотип особи жестко
определяется ее генотипом.
, причем фенотип особи жестко
определяется ее генотипом.
Первоначальная задача исследования состоит в том, чтобы получить
динамическое описание процесса изменения генетической структуры в
череде поколений. Наиболее просто это сделать в случае панмиктичных
популяций, причем при условии, что панмиксия осуществляется при
соединении гамет в зиготы. Обычно делается еще одно допущение.
Рассматриваются диплоидные организмы, у которых половой процесс не
связан с существованием дифференцированных полов, т.е. организмы,
у которых есть диплоидная и гаплоидные стадии, но любые две гаметы
могут образовывать зиготу. Это достаточно сильное ограничение во
многих случаях оказывается эквивалентным более естественному
предположению о том, что начальная генетическая структура по
локусу ![]() одинакова в мужской и женской части популяции [4]
одинакова в мужской и женской части популяции [4]
Введем следующие обозначения: пусть ![]() - концентрация гамет,
несущих аллель
- концентрация гамет,
несущих аллель ![]() ;
; ![]() ,
,![]() ,
,![]() - начальные концентрации
генотипов
- начальные концентрации
генотипов ![]() ,
, ![]() и
и ![]() соответственно;
соответственно; ![]() - численность популяции.
Если нет дифференцированной выживаемости гамет
(т.е. отбор действует только на стадии зигот), то действие отбора
можно описать количественно, поставив в соответствие каждому из
трех генотипических классов особей
- численность популяции.
Если нет дифференцированной выживаемости гамет
(т.е. отбор действует только на стадии зигот), то действие отбора
можно описать количественно, поставив в соответствие каждому из
трех генотипических классов особей ![]() ,
, ![]() и
и ![]() по одному
коэффициенту
по одному
коэффициенту ![]() ,
, ![]() и
и ![]() соответственно, называемому
приспособленностью. Этот коэффициент равен половине числа гамет,
продуцируемых данным генотипическим классом, к численности этого
генотипического класса.
соответственно, называемому
приспособленностью. Этот коэффициент равен половине числа гамет,
продуцируемых данным генотипическим классом, к численности этого
генотипического класса.
Сделанные предположения позволяют получить систему рекуррентных уравнений,
связывающих концентрации генотипических классов гамет и значения
численности популяции в смежных поколениях:
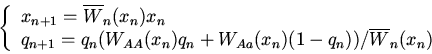
Здесь имеем следующие стационарные точки и условия их устойчивости:
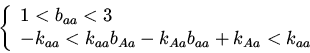
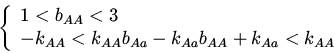
При экспоненциальном экологическом лимитировании функции приспособленности имеют вид:
Для упрощения задачи было введено условие, что существует такая равновесная
точка численности ![]() , в которой приспособленности всех генотипов равны и
приспособленность гетерозиготы постоянна:
, в которой приспособленности всех генотипов равны и
приспособленность гетерозиготы постоянна: ![]() .
Были найдены следующие стационарные точки и условия их устойчивости:
.
Были найдены следующие стационарные точки и условия их устойчивости:
Стационарная точка может существовать при ![]() или
или ![]() , причeм в
первом случае она меньше чем
, причeм в
первом случае она меньше чем ![]() , а во втором - больше
, а во втором - больше ![]() . Устойчива,
если
. Устойчива,
если ![]() ; при
; при ![]() - неустойчива почти всюду.
- неустойчива почти всюду.
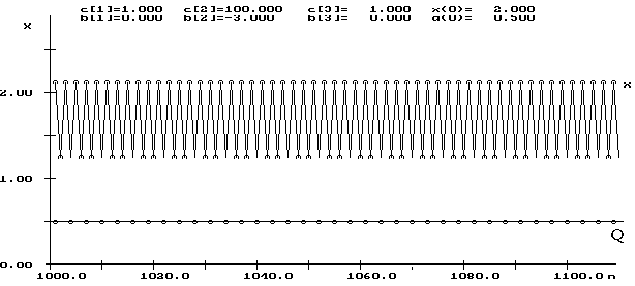
Численное исследование системы в областях потери устойчивости стационарными точками показало, что существуют области параметров, в которых вначале рост средней приспособленности, происходящий в соответствии с фундаментальной теоремой Фишера о естественном отборе, приводит к колебаниям и хаотическим режимам динамики численности. В популяции наблюдается последовательная замена одного доминирующего генотипа другим, а тип наблюдаемого при этом динамического режима численности определяется среднепопуляционным значением мальтузианского параметра. Таким образом, преобладающими оказываются такие режимы эволюции, при которых численность популяции начинает испытывать регулярные или хаотические колебания, а генетический состав выходит на стационарный уровень, причем может наблюдаться как фиксация доминирующих аллеломофов, так и устойчивый (относительно генетических компонентов) полиморфизм (рисунок 1).
 |
В областях, где существуют две неустойчивые стационарные точки, возможны ситуации, когда на фоне флуктуаций численности формируется плотностно-зависимый отбор, и возникают колебания и хаотическая динамика уже генетической структуры популяций (рисунок 2).
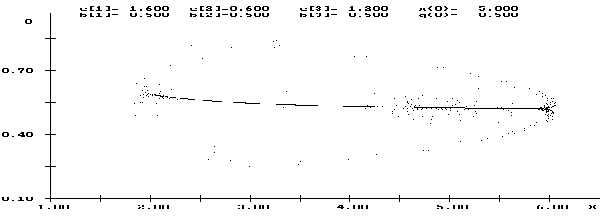 

|
Часто нестационарное поведение численности и генетической структуры
популяции связано с существованием в обобщенном пространстве экологических
и генетических переменных хаотических аттракторов, имеющих сложную
поперечную структуру. Показатели Ляпунова и размерности аттракторов
закономерно меняются с изменением параметров модели (рисунки 3-7).
Проиллюстрируем полученные результаты для случая ![]() ,
, ![]() (экспоненциальная модель). Здесь при
(экспоненциальная модель). Здесь при ![]() (рис. 3) уже хорошо видны
две предельные кривые. При увеличении параметра
(рис. 3) уже хорошо видны
две предельные кривые. При увеличении параметра ![]() очертания предельных
кривых заметно утолщаются. Уже на рисунке 4 видно, что аттрактор имеет
сложную поперечную структуру (по терминологии Хенона). При дальнейшем
увеличении параметра
очертания предельных
кривых заметно утолщаются. Уже на рисунке 4 видно, что аттрактор имеет
сложную поперечную структуру (по терминологии Хенона). При дальнейшем
увеличении параметра ![]() каждая линия аттрактора ``утолщается'' и более
или менее плотно заполняет некоторую область фазового пространства
(рис. 5). Описанную феноменологическую картину можно дополнить
некоторыми количественными характеристиками: оценками значений ляпуновских
показателей (
каждая линия аттрактора ``утолщается'' и более
или менее плотно заполняет некоторую область фазового пространства
(рис. 5). Описанную феноменологическую картину можно дополнить
некоторыми количественными характеристиками: оценками значений ляпуновских
показателей (
![]() , где
, где ![]() - размерность фазового
пространства системы, в данном случае
- размерность фазового
пространства системы, в данном случае ![]() ) и размерности аттрактора
) и размерности аттрактора
![]() (рис. 7). В данном случае вычислялась размерность Каплана - Йорка:
(рис. 7). В данном случае вычислялась размерность Каплана - Йорка:
![]() , где
, где
![]() .
Ляпуновские показатели вычислялись по алгоритму Бенеттина [1].
.
Ляпуновские показатели вычислялись по алгоритму Бенеттина [1].
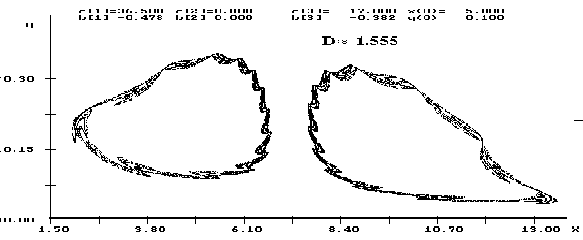
|

|

|

|
На основании полученных результатов можно сказать, что если изменение численности популяции происходит дискретно, в фиксированные периоды размножения, то наличие плотностно зависимого отбора может привести к весьма сложному динамическому поведению численности( а иногда и генетического состава ) даже одной изолированной локальной популяции. В этом случае колебания численности обуславливаются сугубо внутренними процессами в популяции, они не связаны с колебаниями каких-либо внешних факторов. Существуют области параметров популяции, в которых не выполняется гипотеза генетического контроля, так как при значительных колебаниях численности генетический состав популяции остаeтся постоянным, здесь скорее колебания численности расшатывают генетический состав популяции. Колебания же численности популяции в этих областях связаны с ростом еe средней приспособленности [4]. При дальнейшем росте средней приспособленности действительно появляются колебания численности и генетического состава одновременно, что может подтверждать гипотезу генетического контроля.
 Ваши комментарии
Ваши комментарии
|
![[SBRAS]](/gif/s_sigma.gif)
[Головная страница] [Конференции] [СО РАН] |
© 2001, Сибирское отделение Российской академии наук, Новосибирск
© 2001, Объединенный институт информатики СО РАН, Новосибирск
© 2001, Институт вычислительных технологий СО РАН, Новосибирск
© 2001, Институт систем информатики СО РАН, Новосибирск
© 2001, Институт математики СО РАН, Новосибирск
© 2001, Институт цитологии и генетики СО РАН, Новосибирск
© 2001, Институт вычислительной математики и математической геофизики СО РАН, Новосибирск
© 2001, Новосибирский государственный университет
Дата последней модификации Wednesday, 05-Sep-2001 21:00:34 NOVST

