![\begin{displaymath}
x_{n+1}^i=x_{n}^i\exp[R^i(1-\sum_j x_{n}^j/K^i)].\eqno (1)
\end{displaymath}](img2.gif)

Шаповалов А.В.,
Евдокимов Е.В.
Томский государственный университет
Micro-evolution dynamics is studied both analytically and numerically in the framework of the Ricker-May model. The selection is shown to be tended to extension of mean value of population fitness that corresponds to the Fisher's theorem on natural selection in population models with continuous time. The selection in population with a non-zero value of variance in Malthusian and resource parameters is accompanied by successive change of dynamic regimes from a stationary state through limit cycles to chaos and inversely in dependence upon the selection tend. Integrals of motion are presented that results in pairwise combinations of population variables linearly dependent on time. Multidimensional chaotic population dynamics is reduced to a one-dimensional chaotic dynamics.
Ключевые слова: динамика микроэволюционной модели, отбор
Микроэволюционная модель Риккера-Мэя описывает динамику
плотностно-зависимого отбора в биологических популяциях с
бесполым размножением и неперекрывающимися поколениями и имеет
следующий вид [1]:
В работе численными и аналитическими методами исследовались особенности динамики данной системы в случаях, когда в популяции задано такое разнообразие по значениям управляющих параметров, чтобы часть из них находилась в монотонной области, часть - в области предельных циклов, часть - в области хаоса, при этом начальные численности особей разных генотипов существенно отличались.
Показано, что отбор в данной системе всегда направлен в сторону
увеличения средней приспособленности в популяции, определяемой
как
![]() , что
соответствует основной теореме естественного отбора Фишера,
известной для моделей популяций с непрерывным временем. Однако
нелинейные свойства исследуемой системы вносят определенные
особенности в ее эволюционную динамику.
, что
соответствует основной теореме естественного отбора Фишера,
известной для моделей популяций с непрерывным временем. Однако
нелинейные свойства исследуемой системы вносят определенные
особенности в ее эволюционную динамику.
Во-первых, отбор всегда
идет в сторону увеличения ресурсного параметра ![]() , если в
популяциях есть соответствующее генетическое разнообразие. При
его отсутствии отбор как таковой в популяции не наблюдается,
несмотря на наличие дисперсии по мальтузианскому параметру
, если в
популяциях есть соответствующее генетическое разнообразие. При
его отсутствии отбор как таковой в популяции не наблюдается,
несмотря на наличие дисперсии по мальтузианскому параметру ![]() .
.
Во-вторых, в отсутствие дисперсии по ![]() и при наличии
разнообразия по
и при наличии
разнообразия по ![]() в популяции наблюдается сосуществование всех
генотипов. Их равновесное распределение устанавливается в течение
первых трех-четырех поколений и далее не меняется.
в популяции наблюдается сосуществование всех
генотипов. Их равновесное распределение устанавливается в течение
первых трех-четырех поколений и далее не меняется.
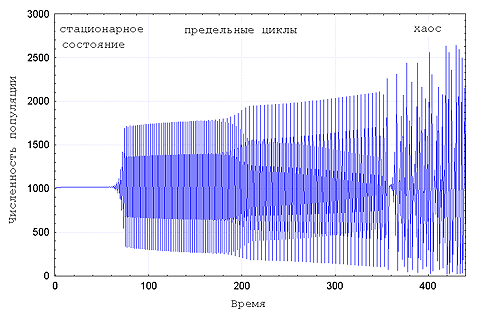
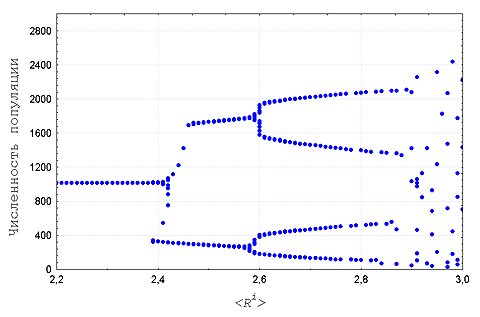
В-третьих, при наличии в популяции разнообразия по
мальтузианскому и ресурсному параметрам течение отбора
сопровождается последовательной сменой динамических режимов от
стационарного состояния через предельные циклы к хаосу и наоборот -
в зависимости от направления отбора по ![]() (Рис. 1).
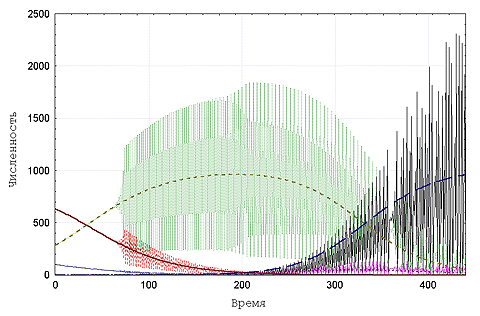
В популяции наблюдается последовательная замена одного
доминирующего генотипа другим, а тип наблюдаемого при этом
динамического режима определяется среднепопуляционным значением
мальтузианского параметра (Рис. 2).
(Рис. 1).
В популяции наблюдается последовательная замена одного
доминирующего генотипа другим, а тип наблюдаемого при этом
динамического режима определяется среднепопуляционным значением
мальтузианского параметра (Рис. 2).
 |
 |
Конкуренция между особями различных генотипов, представленная в
рассматриваемой модели выражением
![]() ,
определяет коллективный тип поведения в системе. Это позволяет
ввести простые парные комбинации популяционных переменных
,
определяет коллективный тип поведения в системе. Это позволяет
ввести простые парные комбинации популяционных переменных
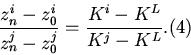
Отображение Риккера-Мэя (1) можно рассматривать как
интегральную форму
![\begin{displaymath}
x^i(t+\delta t)=x^i(t) \exp \left [ R^i\int_t^{t+\delta
t}...
...\displaystyle\frac{\sum_j x^j(\xi)}{K^i}\right )d\xi
\right]
\end{displaymath}](img27.gif)
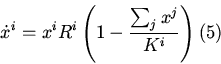
 Ваши комментарии
Ваши комментарии
|
![[SBRAS]](/gif/s_sigma.gif)
[Головная страница] [Конференции] [СО РАН] |
© 2001, Сибирское отделение Российской академии наук, Новосибирск
© 2001, Объединенный институт информатики СО РАН, Новосибирск
© 2001, Институт вычислительных технологий СО РАН, Новосибирск
© 2001, Институт систем информатики СО РАН, Новосибирск
© 2001, Институт математики СО РАН, Новосибирск
© 2001, Институт цитологии и генетики СО РАН, Новосибирск
© 2001, Институт вычислительной математики и математической геофизики СО РАН, Новосибирск
© 2001, Новосибирский государственный университет
Дата последней модификации Sunday, 07-Oct-2001 14:32:48 NOVST