

Чураев Р.Н.
Институт биологии Уфимского научного центра РАН
В представленном докладе показано, как кибернетический подход в трактовке, предложенной А.А. Ляпуновым и С.В. Яблонским (Ляпунов, Яблонский, 1963) и далее более конкретизированной И.А. Полетаевым (Полетаев, 1970), позволяет приблизиться к пониманию таких удивительных биологических феноменов, как наследование приобретенных признаков, частным проявлением которого является хромосомный и геномный импринтинг, а также существование не вписывающихся в дарвиновскую схему эволюционных явлений, например стресс-индуцируемой эволюции и других. Кроме того, в следующем докладе (Галимзянов, 2001) будет показано, что построенный на основе кибернетического подхода простой формализм дает возможность прогнозировать качественную и количественную динамику генных сетей, управляющих жизненными процессами, а также решать задачи структурной, в частности параметрической, устойчивости моделей генных управляющих сетей.
Всякая клетка, в том числе зигота и любая клетка многоклеточного организма, представляет собой систему кодирующих полимеров (ДНК. РНК, белков) и метаболитов - м-систему (Чураев, 1975). Заметим, что во всякой м-системе всегда можно выделить управляющую и управляемую подсистемы, при этом законы динамики этих подсистем различны и описываются для управляющих подсистем дискретными выражениями, а для управляемых - на языке теории дифференциальных уравнений (Чураев, 1986; Tchuraev, 1991).
Таким образом, в клетках можно выделить управляющую систему, в которой как и
в любой управляющей системе можно рассматривать ![]() , представимую как генную сеть над
генетическими элементами (генетическими блоками), включающую в себя
устройства хранения информации ( )
информационные сигнальные процессы
( ), а также
, представимую как генную сеть над
генетическими элементами (генетическими блоками), включающую в себя
устройства хранения информации ( )
информационные сигнальные процессы
( ), а также ![]() молекулярных носителей информации в управляющей генной сети.
Управляющие генные сети (подсети) имеют две основные функции: управление
быстрыми метаболическими процессами синтеза и переработки молекулярных
компонент м-системы, а также управление более медленными онтогенетическими
процессами, приводящими к самовоспроизведению. Далее покажем, как
посредством простого формализма можно исследовать эти функции.
молекулярных носителей информации в управляющей генной сети.
Управляющие генные сети (подсети) имеют две основные функции: управление
быстрыми метаболическими процессами синтеза и переработки молекулярных
компонент м-системы, а также управление более медленными онтогенетическими
процессами, приводящими к самовоспроизведению. Далее покажем, как
посредством простого формализма можно исследовать эти функции.
Пятнадцать лет тому назад был разработан метод обобщенных пороговых моделей
- GTM-формализм для анализа динамики управляющих и управляемых
молекулярно-генетических систем (Чураев, 1986; Tchuraev, 1991). Этот метод,
учитывающий специфику процессов управления на молекулярном уровне, дает
возможность получать как качественные, так и количественные картины динамики
генных сетей произвольной сложности. Основная идея GTM-формализма очень
проста и заключается, как было отмечено выше, в разделении м-системы на
управляющую и управляемую подсистемы, что позволяет большую систему
нелинейных дифференциальных уравнений для молекулярных компонент свести к
кусочно-линейной системе уравнений, правая часть которой содержит
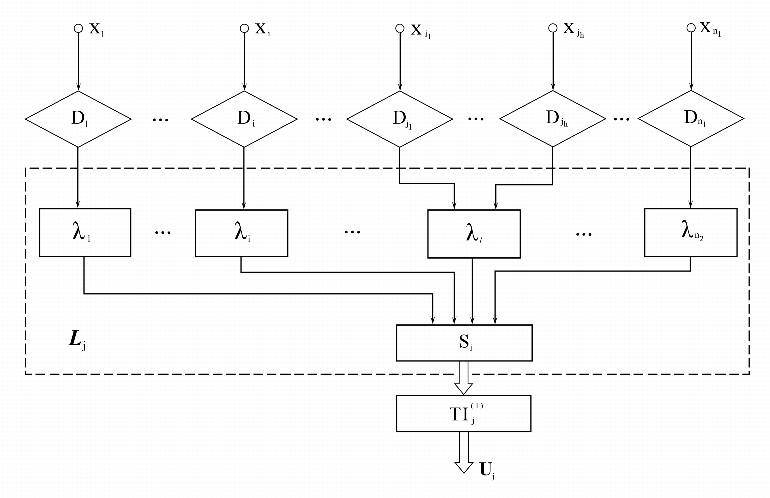
управляющий вектор U![]() = U
= U![]() (t),
формируемый дискретным логическим элементом
(t),
формируемый дискретным логическим элементом ![]() (рис. 1).
(рис. 1).
 |
В случае, когда ![]() -система, то есть молекулярная система кодирующих полимеров и метаболитов, содержит одну копию генома и управление производством
-система, то есть молекулярная система кодирующих полимеров и метаболитов, содержит одну копию генома и управление производством ![]() -го белка
осуществляется на уровне
транскрипции, ранее были получены уравнения динамики следующего вида:
-го белка
осуществляется на уровне
транскрипции, ранее были получены уравнения динамики следующего вида:
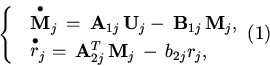
где M![]() , U
, U![]() - столбцы размером
n
- столбцы размером
n![]() 1; A
1; A![]() , B
, B![]() - диагональные матрицы размером n
- диагональные матрицы размером n![]() n
n![]() ;
;
![]() - строка размером 1
- строка размером 1![]() n
n![]() ; r
; r![]() , b
, b![]() - скалярные величины,
причем компоненты
- скалярные величины,
причем компоненты
![]() вектора-столбца
M
вектора-столбца
M![]() обозначают текущую концентрацию, измеренную числом
молекул на клетку, транскриптов
обозначают текущую концентрацию, измеренную числом
молекул на клетку, транскриптов ![]() -й фракции, содержащих j-й цистрон;
U
-й фракции, содержащих j-й цистрон;
U![]() = U
= U![]() (t) - управляющий
вектор, формируемый логическим элементом и элементами задержки данного
блока; диагональный элемент
(t) - управляющий
вектор, формируемый логическим элементом и элементами задержки данного
блока; диагональный элемент
![]() матрицы A
матрицы A![]() обозначает единичную интенсивность
транскрипции, начинающейся с одной копии
обозначает единичную интенсивность
транскрипции, начинающейся с одной копии ![]() -го промотора; соответственно
компонента
-го промотора; соответственно
компонента
![]() строки
строки
![]() - единичная интенсивность трансляции с
- единичная интенсивность трансляции с ![]() -го
транскрипта; диагональный элемент
-го
транскрипта; диагональный элемент
![]() матрицы B
матрицы B![]() - единичная интенсивность
(коэффициент) деградации
- единичная интенсивность
(коэффициент) деградации ![]() -го транскрипта; b
-го транскрипта; b![]() - коэффициент деградации j-го полипептида.
- коэффициент деградации j-го полипептида.
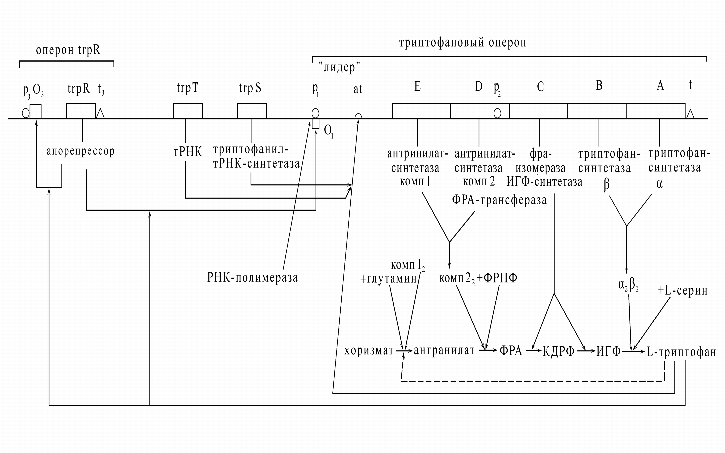
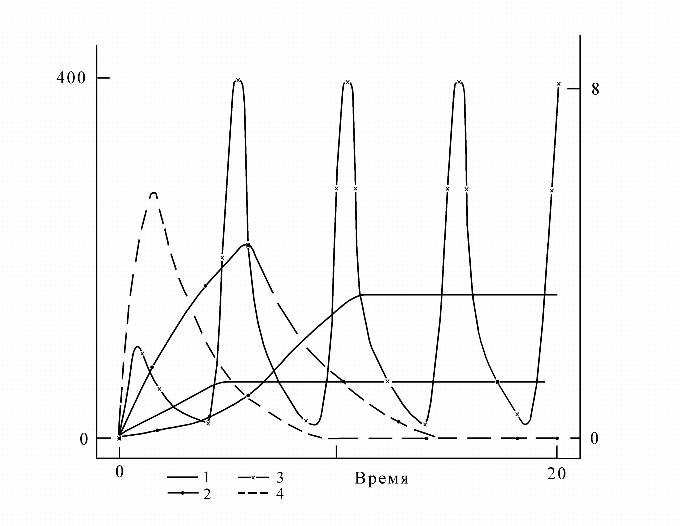
Далее приведем пример результата, полученного на построенной этим методом модели системы управления анаболическим процессом синтеза триптофана. На рис. 2 представлена система управления синтезом аминокислоты триптофана.
 |
 |
Методом обобщенных пороговых моделей также была построена и исследована математическая модель системы управления катаболизмом арабинозы (Prokudina et al., 1991).
Посредством частной версии GTM-формализма в 70-е годы были построены первые
две модели системы управления развитием ![]() -фага: СУРФ-
-фага: СУРФ-![]() и
СУРФ-
и
СУРФ-![]() (Чураев, Ратнер, 1975; Ratner, Tchuraev, 1978; Kananyan et
al., 1981). Даже столь простой объект как
(Чураев, Ратнер, 1975; Ratner, Tchuraev, 1978; Kananyan et
al., 1981). Даже столь простой объект как ![]() -фаг, геном которого
содержит всего 48 генов, имеет сложную управляющую генную сеть (рис. 4).
-фаг, геном которого
содержит всего 48 генов, имеет сложную управляющую генную сеть (рис. 4).
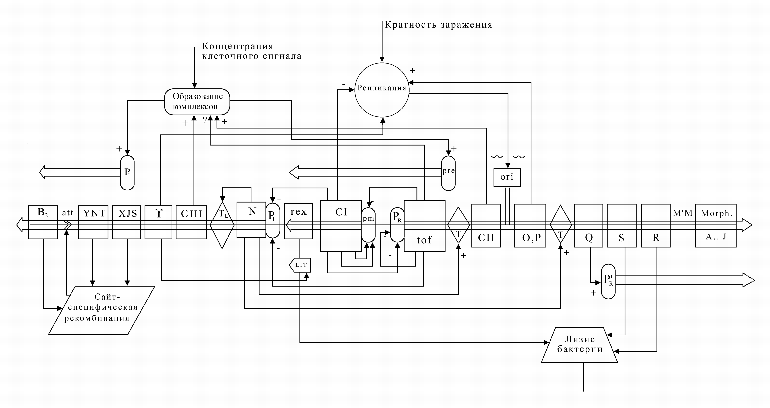 |
На следующем рисунке (рис. 5) представлены кинетические кривые для транскриптов в лизогенном и литическом режимах.
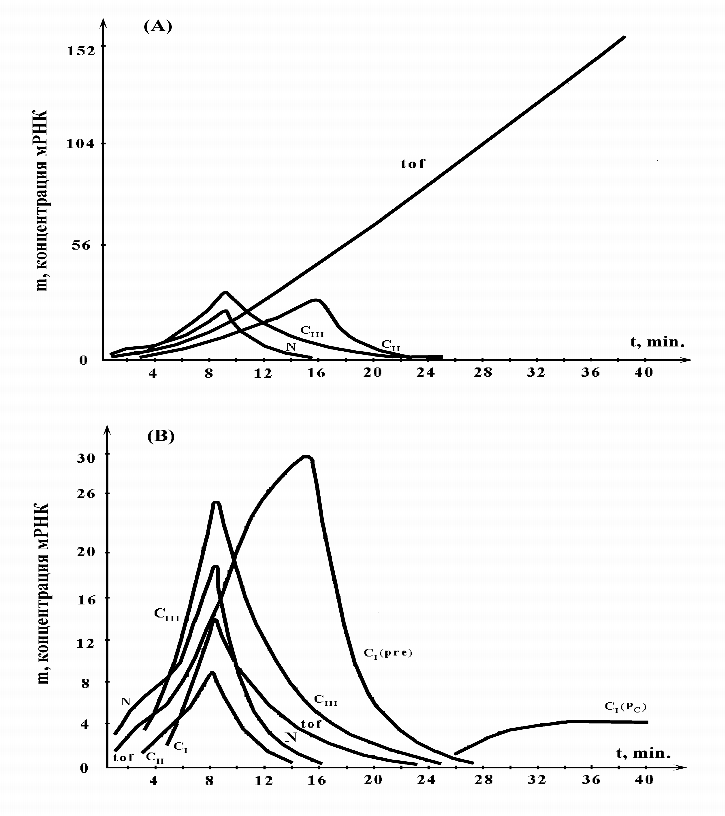 |
С учетом этих особенностей уравнения динамики имеют следующий вид:
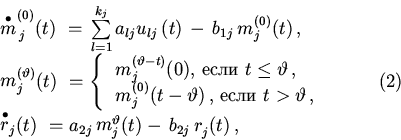
Посредством сопутствующего пакета программ (Galimzyanov, 2000) нам удалось построить и получить кинетические кривые, а также исследовать на параметрическую устойчивость две модели эукариотических генных сетей: системы управления морфогенезом цветка Arabidopsis thaliana и системы управления ранним онтогенезом у Drosophila (Чураев, Галимзянов, 2001). Результаты представлены в следующем докладе (Галимзянов, 2001).
Как уже было отмечено, в молекулярных системах кодирующих полимеров и метаболитов всегда можно вычленить управляющие подсистемы, в которых протекают сигнальные процессы, управляющие биохимическими процессами производства молекулярных компонент системы. Среди этих управляющих подсистем особый интерес представляют системы управления онтогенезом (СУО). Некоторые из этих систем изучены как экспериментально, так и теоретически. В системах управления онтогенезом происходят процессы переработки информации, как содержащейся в самой системе, так и поступающей извне. Если не вдаваться в детали строения OCS, то можно ее рассматривать как "черный ящик", имеющий конечное число входных и выходных каналов.
На языке теории автоматов была построена в общем виде модель системы
управления онтогенезом (автомат A![]() (Чураев, 1997;
Tchuraev, 2000). Выбор математического аппарата именно этой теории диктуется
тем, что изучаемые нами процессы управления по сути своей дискретны, а
автоматы моделируют устройства переработки информации.
(Чураев, 1997;
Tchuraev, 2000). Выбор математического аппарата именно этой теории диктуется
тем, что изучаемые нами процессы управления по сути своей дискретны, а
автоматы моделируют устройства переработки информации.
Было показано, что если во множестве ![]() состояний памяти
состояний памяти ![]() автомата
автомата ![]() существует по крайней мере два
различимых возвратных состояний
существует по крайней мере два
различимых возвратных состояний
![]() и
и
![]() а среди всевозможных входных
последовательностей такая последовательность
а среди всевозможных входных
последовательностей такая последовательность
![]() , что приложение ее к
, что приложение ее к
![]() /
/
![]() либо к
либо к ![]() /
/
![]() приведет к состояниям
приведет к состояниям
![]() и
и
![]() соответственно, то эти состояния наследуются, то есть
состояние
соответственно, то эти состояния наследуются, то есть
состояние
![]() памяти
памяти
![]() автомата
автомата ![]() зависит от состояния памяти
зависит от состояния памяти
![]() , где
, где ![]() - соответствует
номеру клеточного поколения. Также было показано, что если нетривиальный
неавтономный автомат
- соответствует
номеру клеточного поколения. Также было показано, что если нетривиальный
неавтономный автомат
![]() содержит по
крайней мере одну элементарную ячейку памяти, которая имеет два состояния,
взаимно-однозначно соответствующие двум альтернативным подпрограммам
онтогенеза, то эти состояния наследуемые (Чураев, 1997).
содержит по
крайней мере одну элементарную ячейку памяти, которая имеет два состояния,
взаимно-однозначно соответствующие двум альтернативным подпрограммам
онтогенеза, то эти состояния наследуемые (Чураев, 1997).
При интерпретации результатов математического моделирования систем
управления онтогенезом следует иметь ввиду, что автоматы ![]() (модели СУО) чисто функциональные конструкции, поскольку их
внутренняя структура не рассматривается: в этих автоматах состояния
функциональной памяти всего лишь "внутренние переменные", позволяющие
связывать выходные переменные автомата со входными, то есть нет "привязки" к
реальным структурам генных систем. Вследствие этого функции переходов (
(модели СУО) чисто функциональные конструкции, поскольку их
внутренняя структура не рассматривается: в этих автоматах состояния
функциональной памяти всего лишь "внутренние переменные", позволяющие
связывать выходные переменные автомата со входными, то есть нет "привязки" к
реальным структурам генных систем. Вследствие этого функции переходов (![]() и выходов (
и выходов (![]() ) могут быть одинаковыми для совершенно по-разному
устроенных молекулярных механизмов, их реализующих. Поэтому в этом смысле
можно говорить о функциональной эквивалентности как систем управления онтогенезом, моделируемых одним
классом автоматов, так и молекулярных механизмов, реализующих функциональную
память СУО, в частности функциональную наследственную память.
) могут быть одинаковыми для совершенно по-разному
устроенных молекулярных механизмов, их реализующих. Поэтому в этом смысле
можно говорить о функциональной эквивалентности как систем управления онтогенезом, моделируемых одним
классом автоматов, так и молекулярных механизмов, реализующих функциональную
память СУО, в частности функциональную наследственную память.
Таким образом, функциональная наследственная память как категория - множество функционально эквивалентных молекулярных механизмов, обеспечивающих кодирование, хранение и передачу в ряду генераций функциональной наследственной информации.
Интерпретация: существование эпигенов. Что является материальным воплощением ячеек функциональной наследственной
памяти? Много лет тому назад была предложена гипотеза о существовании
особого класса наследственных единиц - ![]() , в которой часть наследственной
информации хранится, кодируется и передается потомству вне молекул ДНК
генома (Чураев, 1975; Tchuraev, 1997). Тогда в качестве моделей эпигенов
были рассмотрены лишь циклические системы генов. Но, если не ограничиваться
только циклическими генными системами, то эпигеном можно называть каждую систему
генов, имеющую не менее двух устойчивых режимов фунционирования подчиненных
ей генов и способную сохранять каждый из режимов в последовательном ряду
генераций. Надо сказать, что к настоящему времени накопилось большое число
фактов, свидетельствующих в пользу этой гипотезы. Более того, появилась
ветвь биологии - эпигенетика, суть предмета которой, по определению А. Риггса,
сделанному пять лет тому назад, такова: ``Эпигенетика - изучение митотически
и/или мейотически наследуемых изменений в функции гена, которые не могут
быть объяснены изменениями в последовательности ДНК'' (Riggs, 1996).
, в которой часть наследственной
информации хранится, кодируется и передается потомству вне молекул ДНК
генома (Чураев, 1975; Tchuraev, 1997). Тогда в качестве моделей эпигенов
были рассмотрены лишь циклические системы генов. Но, если не ограничиваться
только циклическими генными системами, то эпигеном можно называть каждую систему
генов, имеющую не менее двух устойчивых режимов фунционирования подчиненных
ей генов и способную сохранять каждый из режимов в последовательном ряду
генераций. Надо сказать, что к настоящему времени накопилось большое число
фактов, свидетельствующих в пользу этой гипотезы. Более того, появилась
ветвь биологии - эпигенетика, суть предмета которой, по определению А. Риггса,
сделанному пять лет тому назад, такова: ``Эпигенетика - изучение митотически
и/или мейотически наследуемых изменений в функции гена, которые не могут
быть объяснены изменениями в последовательности ДНК'' (Riggs, 1996).
Поскольку, согласно определению, эпигены способны сохранять функциональные состояния (или последовательности функциональных состояний, то есть режимов) в ходе онтогенеза и обеспечивать передачу кодируемой этими состояниями функциональной информации, естественно полагать, что молекулярно-генетической реализацией ячеек функциональной наследственной памяти являются эпигены.
Приведем пример реально существующего простейшего эпигена у ![]() -фага
(рис. 6).
-фага
(рис. 6).
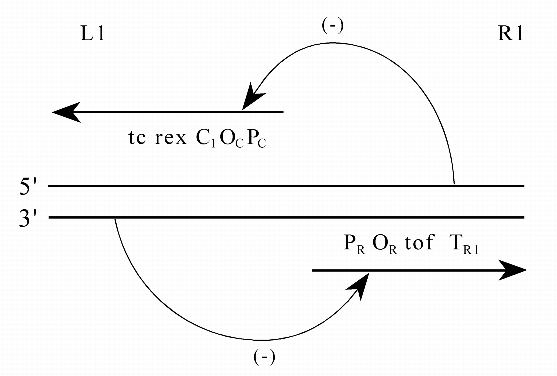 |
Заметим, что эпигены - надгенные системы, то есть "сделаны" из генов или их
частей. Таким образом, ![]() и
и ![]() - наследственные единицы разных рангов.
- наследственные единицы разных рангов.
В большом обзоре известного генетика Отто Ландмана (1991) приведено много фактов, реанимирующих казалось бы давно закрытую проблему наследования приобретенных признаков. Легко видеть, что эпигенные системы, содержащие эпигены, способны обеспечивать наблюдаемые феномены наследования приобретенных признаков.
А теперь перейдем к рассмотрению роли эпигенетических систем в филогенезе. В эволюционном плане существование эпигенов, так сказать, второго, дополнительного потока наследственной информации, приводит к интересным следствиям. Упомянем некоторые из них. Изменчивость, обусловленная эпигенами, может, в отличие от ненаправленной генной изменчивости, быть связана с состоянием среды. Можно сказать, что эпигены и эпигенные системы упорядочивают общую наследственную случайную изменчивость и тем самым до определенной степени канализируют процесс эволюции. Таким образом, эпигенная изменчивость не носит, как правило, случайный характер и может привести к формированию из готовых блоков (подпрограмм) новых вариантов наследственных программ онтогенеза по принципу тинкеринга, то есть когда каждый предыдущий блок может функционировать как целое. Существование эпигенов, функциональной наследственной памяти, по-видимому, дает возможность для реализации другой (недарвиновской) эволюционной стратегии, когда относительно неудачные эволюционные ходы в наследственной памяти некоторых особей "не забываются", и соответствующие подпрограммы сохраняются в выключенном состоянии в функциональной наследственной памяти, не проявляясь в онтогенезе (Чураев, 1987; Tchuraev, 2000).
Теперь хотелось бы обратиться к прикладным аспектам исследования феномена наследственности. Без малого тридцать лет развивается генная инженерия, методы которой позволяют манипулировать частями геномов, выделять и переносить определенные гены одного организма к другому, синтезировать искусственные гены, если известны их первичные нуклеотидные последовательности. Уже давно никого не удивить трансгенными продуктами.
Однако эти замечательные исследования имеют некий ``потолок'', который определяется тем, что, во-первых, неизвестно, каким образом функция гена закодирована в последовательности его нуклеотидов, во-вторых, заданное структурное преобразование выбранной последовательности нуклеотидов представляет собой исключительно трудную техническую задачу. Это приводит к тому, что на пути преобразования структуры генома чрезвычайно сложно получить качественно новую заданную наследственную информацию, качественно новый требуемый признак или комплекс признаков. Иными словами, основная проблема прикладной генетики - управление наследственной информацией организмов - может быть разрешена методами генной инженерии в некоторой области, ограниченной манипулированием уже существующими методами. Нет сомнения, что с развитием генетической инженерии границы этой области будут расширяться.
Довольно давно автором был предложен иной подход к проблеме управления наследственной информацией, который основан на конструировании методами генной инженерии эпигенов с наперед заданными управляемыми наследуемыми функциональными свойствами. Теоретически была показана возможность конструирования любого эпигена с наперед заданным разнообразием наследуемых признаков. Заметим, что основной особенностью эпигенов является возможность целенаправленного изменения содержащейся в них части наследственной информации без изменения нуклеотидной последовательности генома. (Чураев, 1982).
Таким образом, идея эпигенной инженерии заключается в том, что из генов и их частей
конструируются эпигенные системы (системы следующего ранга сложности, чем
гены) с прогнозируемыми заданными управляемыми наследуемыми режимами
функционирования входящих в них генов. Для того чтобы проверить эту идею, в
нашей лаборатории были проведены несложные, но очень трудоемкие
эксперименты. В результате, впервые параллельно и независимо от группы
Гарднера (Масaчусетский университет, США) были получены циклические дигенные
плазмидные конструкции, имеющие два устойчивых альтернативных функциональных
состояния, пример одной из которых приведен на рис.7 (Tchuraev at al., 2000;
Gardner et al., 2000). Мы экспериментально показали, что эта система
является элементарным ![]() , т.е. может кодировать, хранить и передавать
клеточному потомству (наследовать) вызванные внешними специфическими
сигналами изменения функционального состояния без изменения ДНК-последовательностей, входящих в систему генов.
, т.е. может кодировать, хранить и передавать
клеточному потомству (наследовать) вызванные внешними специфическими
сигналами изменения функционального состояния без изменения ДНК-последовательностей, входящих в систему генов.
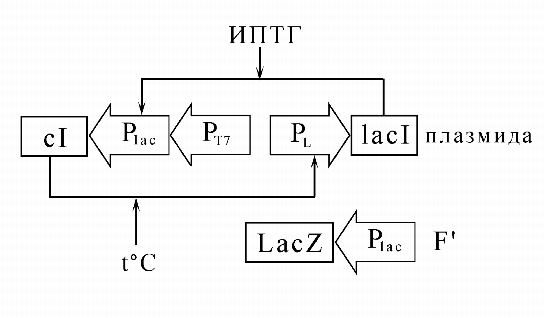 |
На рис. 8 показано наследование функциональных состояний.
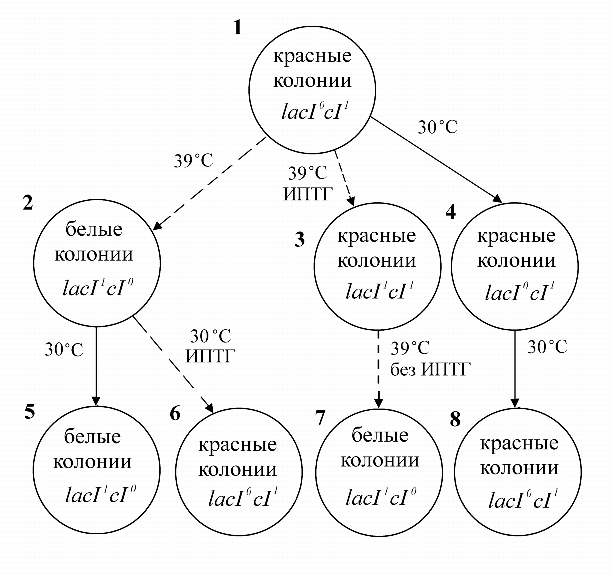 |
Полученный нами простой эпиген по существу представляет ячейку памяти, способную ``запоминать'' двоичные сигналы, подобно элементарным ячейкам запоминающих устройств компьютеров. Надо однако заметить, что наши ячейки имеют субмолекулярные размеры и помещаются в бактериях, объемом в один кубический микрон. Ясно, что можно последовательно ``наращивать'' такие простые искусственные генные сети с тем, чтобы они реализовывали все более сложные функции. Такие синтетические генные сети могут найти широкое применение в биотехнологии, например для управления динамикой размножения клеточных культур - продуктов биологически активных веществ (инсулина, соматостатина и др.). Возможно их применение и в генной терапии - для адресного включения компенсаторных генов в нужный момент онтогенеза при лечении наследственных заболеваний.
Приведем пример одной молекулярно-генетической конструкции, реализующий штрих Шеффера (рис. 9). Поскольку функции Шеффера образуют полную систему функций, то, конструируя такие блоки, можно реализовать любую булеву функцию. Возможно также конструировать из генов системы, реализующие арифметические операции по модулю 2.
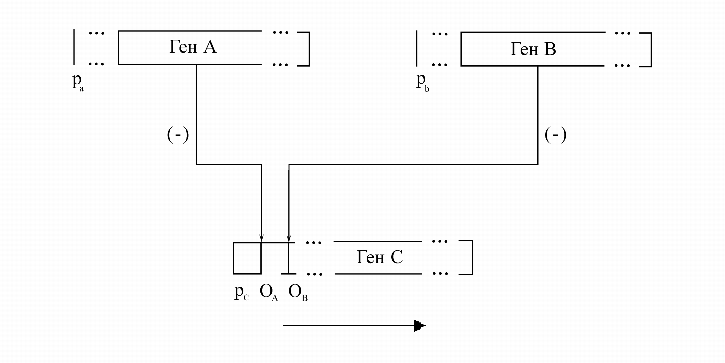 |
Таким образом, в принципе можно, манипулируя генами и их частями, получать
молекулярные устройства, реализующие логические и арифметические операции, а
так же ячейки памяти, то есть молекулярные микросхемы - ![]() . Поскольку
ДНК-носители этих микросхем автоматически воспроизводятся универсальными
клеточными устройствами редупликации, то сконструированный молекулярный
внутриклеточный ``чип'' будет тиражироваться, причем и, как показано выше,
функциональные состояния ячеек памяти будут сохраняться. Так что, наверное,
можно в перспективе конструировать молекулярно-генетические биокомпьютеры.
. Поскольку
ДНК-носители этих микросхем автоматически воспроизводятся универсальными
клеточными устройствами редупликации, то сконструированный молекулярный
внутриклеточный ``чип'' будет тиражироваться, причем и, как показано выше,
функциональные состояния ячеек памяти будут сохраняться. Так что, наверное,
можно в перспективе конструировать молекулярно-генетические биокомпьютеры.
Всякая живая система с очевидностью должна обладать свойством устойчивости в широком смысле, обеспечивающей процесс самовоспроизведения. В частности, и онтогенез как совокупность внутрисистемных процессов, приводящих к самовоспроизведению живых систем, должен быть устойчивым как к внешним воздействиям, так и к внутренним флуктуациям. Устойчивость онтогенеза в значительной степени обеспечивается системой, управляющей онтогенезом
В следующем докладе будет рассказано о математических моделях конкретных подсистем управления онтогенезом. Как отметил В.И.Арнольд (1997), выводам, сделанным на основании исследования жестких математических моделей реальных систем, можно доверять лишь тогда, когда они подтверждаются исследованием их структурной устойчивости, в частности параметрической. Если известно, что моделируемый объект устойчив в широком смысле, то и его адекватная модель должна обладать структурной устойчивостью. Поэтому в сопутствующем докладе будут приведены результаты исследования этих моделей на структурную (параметрическую) устойчивость.
Согласно А.А.Ляпунову ``жизнь можно охарактеризовать как высокоустойчивое состояние вещества, использующее для выработки сохраняющих реакций информацию, кодируемую состояниями отдельных молекул'' (Ляпунов, 1963). ``Состояния отдельных молекул'' -
это в современной терминологии структурные состояния частей молекул ДНК,
имеющих начало и конец (единиц транскрипции - генов). Различные структурные
состояния генов - это различные 4-х буквенные слова (А, Г, Ц, Т) конечной
длины, причем информация в генах кодируется структурным способом. Однако,
как было показано выше, для выработки сохраняющих реакций, то есть
внутренних реакций вещества на внешние воздействия, направленных на
сохранение своего состояния, используется и информация, кодируемая
функциональными состояниями молекулярной системы кодирующих полимеров и
метаболитов. Поэтому можно сказать, что ![]() процесс, характеризующийся высокоустойчивыми состояниями вещества, организованного в многоярусные системы кодирующих полимеров и метаболитов высокой сложности, использующего для выработки сохраняющих реакций как структурную информацию, кодируемую структурными состояниями отдельных молекул (кодирующих молекул), так и функциональную информацию, кодируемую функциональными состояниями м-систем.
процесс, характеризующийся высокоустойчивыми состояниями вещества, организованного в многоярусные системы кодирующих полимеров и метаболитов высокой сложности, использующего для выработки сохраняющих реакций как структурную информацию, кодируемую структурными состояниями отдельных молекул (кодирующих молекул), так и функциональную информацию, кодируемую функциональными состояниями м-систем.
Таким образом, кибернетический (информационно-операционный) подход к изучению явлений живой природы, на мой взгляд, позволяет свести в единую систему внешне противоречивые как давно известные, так и новейшие данные, полученные в рамках и классической, и современной биологии.
Автор глубоко благодарен Е.Л.Пущину за плодотворные дискуссии и А.В.Галимзянову за техническую помощь в подготовке рукописи.
 Ваши комментарии
Ваши комментарии
|
![[SBRAS]](/gif/s_sigma.gif)
[Головная страница] [Конференции] [СО РАН] |
© 2001, Сибирское отделение Российской академии наук, Новосибирск
© 2001, Объединенный институт информатики СО РАН, Новосибирск
© 2001, Институт вычислительных технологий СО РАН, Новосибирск
© 2001, Институт систем информатики СО РАН, Новосибирск
© 2001, Институт математики СО РАН, Новосибирск
© 2001, Институт цитологии и генетики СО РАН, Новосибирск
© 2001, Институт вычислительной математики и математической геофизики СО РАН, Новосибирск
© 2001, Новосибирский государственный университет
Дата последней модификации Monday, 10-Sep-2001 20:48:33 NOVST
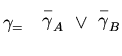 , где бинарная переменная
, где бинарная переменная