Анализ расщепления гибридов методом главных компонент 1
Маркель А.Л. Лаборатория эволюционной генетики, Институт цитологии и генетики СО РАН Ефимов В.М. Лаборатория зоомониторинга, Институт систематики и экологии животных СО РАН Ковалева В.Ю.
Лаборатория охраны генофонда, Институт систематики и экологии животных СО РАН
Аннотация:
Методом главных компонент проанализирована объединенная внутривыборочная
изменчивость лабораторных крыс двух чистых линий - НИСАГ
(ISIAH) и WAG - и первых двух
поколений гибридов между ними, у которых промерены 20
морфологических, физиологических и поведенческих признаков. На первые пять
компонент пришлось 64.4% общей дисперсии. Первую и третью
компоненты образовали все поведенческие признаки, за исключением величины
дефекации и латентного периода, вторую - веса внутренних органов и, в
противофазе с ними, базальное артериальное давление. По второй компоненте
проявился гетерозис. Наиболее интересными оказались IV и V
компоненты, в которые вошли все признаки, характеризующие стрессированность
животных. Выявлена их линейная комбинация, наибольший вклад в которую вносит
артериальное давление при стрессе и наследуемость которой соответствует
аддитивной модели без доминирования
(h=0.51; F=2.1; p<0.05).
Joint intra-sample diversity of laboratory rats samples from two pure lines
- ISIAH and WAG - and two first hybrid generations was
analyzed by principal components method. Twenty morphological, physiological
and behavioral traits were used. First five components occupied
64.4% of common variance. All behavioral traits with the
exception defecation level and latent interval formed first and third
components, all interior organs masses vs basal arterial pressure formed
second component. On this component heterosis was appear. Fourth and fifth
components proved of most interest, because all traits characterized animal
stress mainly arterial pressure in stress formed them. A linear combination
of "stress'' components was found, which inheritance corresponded to
additive model without dominance (h=0.51;
F=2.1; p<0.05).
Ключевые слова: расщепление гибридов, метод главных компонент.
Еще в 1941 году Алексей Андреевич Ляпунов совместно с Ю.Я.Керкисом
опубликовал статью "О расщеплении гибридов''[1], посвященную
статистическому анализу результатов генетических экспериментов по
скрещиванию. Работа была выполнена по инициативе Н.И.Вавилова и
А.Н.Колмогорова. Современное возрастание мощности вычислительных машин,
основоположником и активным пропагандистом внедрения которых в биологию
являлся А.А.Ляпунов, сделало возможным, помимо многого другого, практическое
применение к этим классическим областям генетики многомерного
статистического анализа.
Хорошо известно, что фенотипическая изменчивость гибридов
F от скрещивания двух чистых линий является
ненаследственной и только начиная с F в
изменчивости проявляется расщепление комплексов генов, полученных от обоих
родителей. Если мы анализируем одновременно несколько количественных
признаков, на которые совместно действует совокупность многих генов и
становление которых происходит под действием общих онтогенетических
механизмов, то онтогенетическая и генетическая общность этих признаков
фенотипически должны проявляться в коррелятивных связях между ними. И
наоборот, распад коррелятивных связей на несколько статистически независимых
подсистем является указанием на их относительную онто- и генетическую
независимость. Метод главных компонент позволяет разложить общую
фенотипическую изменчивость на несколько ортогональных составляющих, которые
являются новыми признаками и могут анализироваться любыми одномерными и
многомерными методами генетического анализа.
Этим методом проанализирована изменчивость выборок лабораторных крыс двух
чистых линий - НИСАГ (ISIAH) и
WAG - и первых двух поколений гибридов между ними, у
которых промерены 20 морфологических, физиологических и
поведенческих признаков. Для нормализации распределения все признаки, за
исключением артериального давления, подвергались логарифмическому
преобразованию [3,5]. Считается, что родительские поколения и первое
поколение гибридов генетически однородны и между собой различаются только
средними значениями вследствие разницы в генных наборах, а наблюдаемая
фенотипическая изменчивость внутри каждой выборки целиком обусловлена
стохастическими отклонениями в ходе онтогенеза. Эта изменчивость
структурируется программой развития особи, обеспечивающей целостность
организма [4]. Во втором поколении к упорядоченной онтогенезом многомерной
изменчивости добавляется фенотипическое разнообразие, вызванное генетическим
расщеплением. Чтобы выявить ее структурированность, каждая выборка
центрировалась отдельно, после чего они были объединены и были вычислены
первые пять компонент объединенной внутривыборочной изменчивости (таб.1)
[2].
Т а б л и ц а 1
Коэффициенты корреляции (x1000)
морфологических,
физиологических и поведенческих признаков
с первыми пятью
главными компонентами
объединенной внутривыборочной изменчивости
N
Признаки
Компоненты
I
II
III
IV
V
Ab
Базальное артериальное давление
-049
-440***
-068
488***
-078
As
Артериальное давление
при стрессе
-103
-059
-046
685***
-312***
Pb
Вес тела
-127
907***
111
-015
030
Pa
Вес надпочечников
-318***
400***
052
447***
-042
Pk
Вес почек
-161
781***
185*
024
-134
Pc
Вес сердца
-322***
768***
-059
130
047
Ob
Базальная концентрация кортикостерона
в плазме крови
-239**
-083
-069
091
531***
Os
Концентрация кортикостерона
в плазме крови
при стрессе
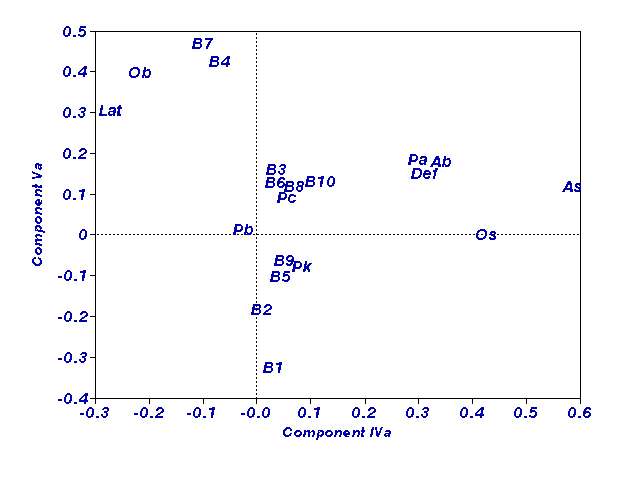
Рис. 1.
Расположение признаков на плоскости компонент IVa и Va
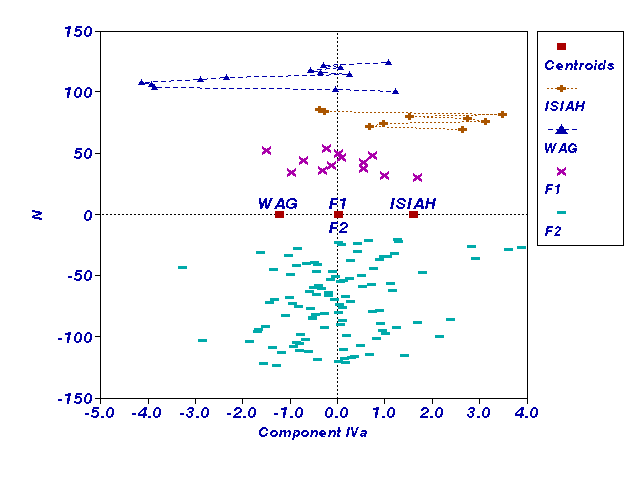
Рис. 2.
Соответствие распределения выборок по компоненте IVa
аддитивной модели наследования без доминирования (ось ординат - номера
особей, у F2 - с минусом).
На первые пять компонент пришлось 64.4% дисперсии. Первую и
третью компоненты, в основном, образовали поведенческие признаки, вторую -
веса внутренних органов и, в противофазе с ними, базальное артериальное
давление. По второй компоненте проявился гетерозис. Наиболее интересными
оказались IV и V компоненты, в которые вошли все признаки,
характеризующие стрессированность животных. Выявлена их линейная комбинация,
наибольший вклад в которую вносит артериальное давление при стрессе (рис.1)
и наследуемость которой соответствует аддитивной модели без доминирования
(h=0.51; F=2.1; p<0.05) (рис. 2,
ось ординат введена для удобства).


![[SBRAS]](/gif/s_sigma.gif)